Animals large and small use muscle to move – but can they all achieve the same relative performance? We combine results from dimensional analyses with empirical data on muscle physiology and anatomy to explain (and ideally predict!) the variation of animal performance with size. By linking physical models with animal size, we hope to identify opportunities for non-trivial musculoskeletal adaptations that distinguish animals across body sizes and locomotor mode.

Striated muscle looks more or less the same, whether an animal swims, runs, crawls, or flies; whether it is smaller than the tip of a sharp pencil or heavier than 15 school buses; whether it first appeared millions or only thousands of years ago. Does this shared design have shared implications? Starting from first principles, we seek to identify dimensionless numbers that enable fair comparison of muscle performance across animal size, physiological make-up, and environmental setting.
Read A theory of physiological similarity in muscle-driven motion

Insect mandibles are “multi-purpose” tools: they are used for cutting, masticating, digging, material transport, prey capture and defense. In contrast to this functional complexity, the kinematic space of most insect mandibles is thought to be fully characterised by a simple hinge joint. Although widespread, this assumption is based solely on morphological inference; quantitative kinematic analysis are lacking.
We developed a recording setup and kinematic analysis framework to rigorously test the “hinge joint hypothesis” in leaf-cutter ants. At large opening angles, mandibles yaw, consistent with a simple hinge joint. But at small opening angles, mandibles can both yaw and pitch, suggesting multiple degrees of freedom, and that insect mandibles may be capable of more complex motion than traditionally assumed.

Insects seem to move effortlessly in a variety of habitats. They can walk upright, climb up vertical walls, stick to inverted surfaces, and are robust to the loss of one or even multiple limbs. We track points of interest using markerless pose estimation and reconstruct the data in 3D for further mechanical and statistical analysis. Why do insects move the way they do?

How well can insects cling to surfaces of different properties?
On rough surfaces, such as rocks or tree bark, insects can employ not only their pads but also their claws to help them attach. On smooth surfaces, however, claws become useless, and insects have to rely on intermolecular forces. What role does surface chemistry play in the pad-surface interaction? Variations in surface chemistry lead to differences in the ability of contacting liquids to spread – different surface chemistry usually implies different wettability. To test whether surface chemistry influences adhesion, we produce smooth surfaces of varying wettability by Chemical Vapour Deposition (CVD), characterise them by goniometry and Atomic Force Microscopy, and perform single pad adhesion force measurements using a custom-made fibre-optic force sensor.

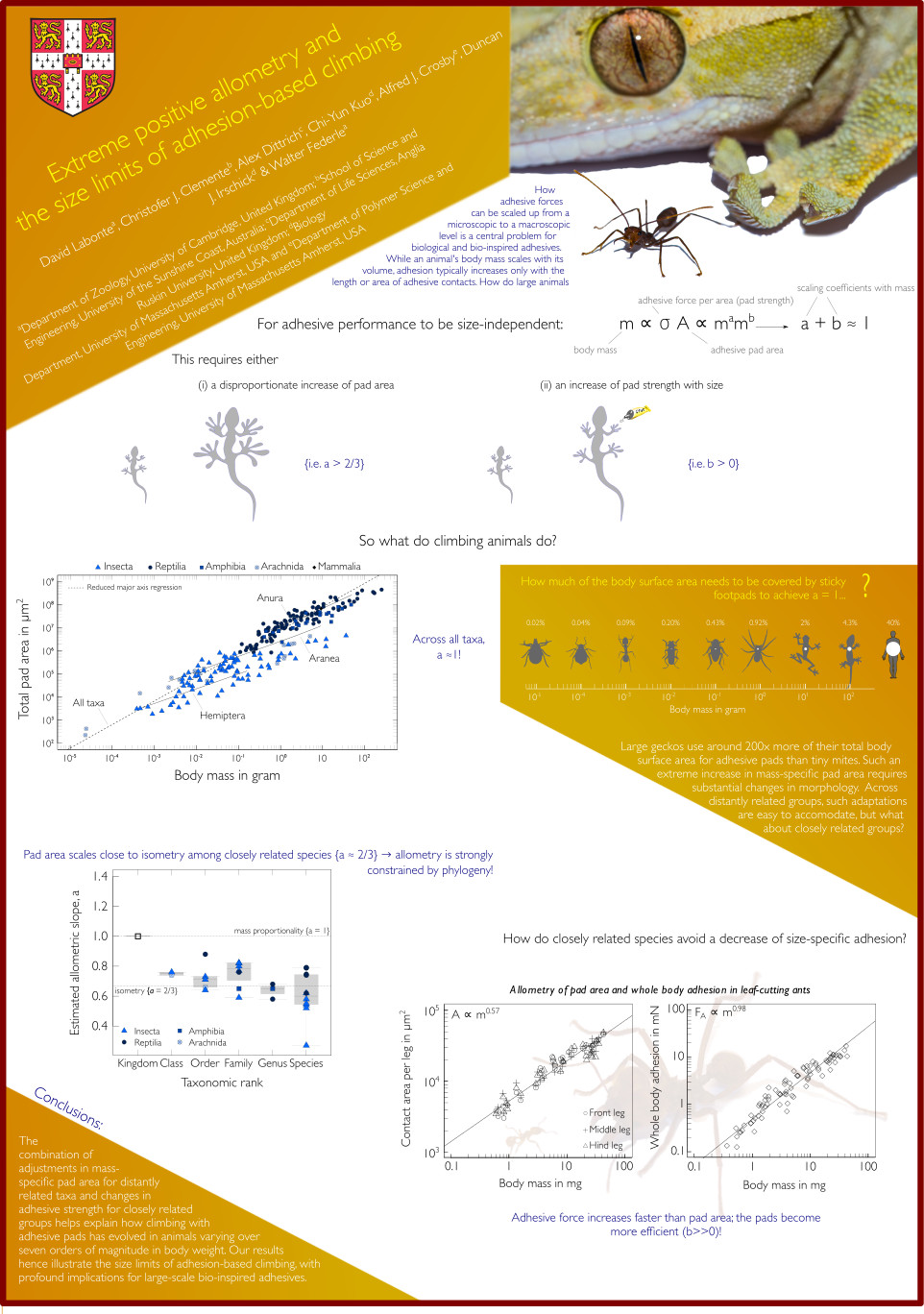
How do climbing animals safely attach to surfaces across large difference in body size?
Textbook scaling theory predicts that larger animals are increasingly challenged: the force that adhesive patches needs to sustain depends on body volume, but the sticky area depends on body surface area. One way to overcome this problem would be a disproportionate increase in pad area. However, animal footpads only become disproportionally larger across very distant groups; within closely related clades, they appear to become more sticky instead! What is the source of this enhanced efficiency? We explore this problem theoretically, and through experimental work which focusses on the physical properties of the insect pad secretion.
We test whether the surface tension and viscosity of the secretion vary systematically with size, by combining Interference Reflection Microscopy and dewetting measurements on individuals spanning almost 3 orders of magnitude in mass. Learning how natural adhesive organs overcome stress concentrations and scaling issues to maintain their attachment performance may inform the next generation of artificial adhesives.
Read: Shear-sensitive adhesion enables size-independent adhesive performance in stick insects | The physical properties of the stick insect pad secretion are independent of body size